20/04/2024 às 09h04min - Atualizada em 20/04/2024 às 09h04min
Formação de biofilme e polissacarídeos capsulares são estratégias inteligentes que Staphylococcus aureus utiliza para sobreviver e causar doenças
O presente estudo oferece insights valiosos sobre a caracterização fenotípica e molecular do Staphylococcus aureus em contextos de mastite bovina
Alessandra Pereira Sant’Anna Salimena, Letícia Scafutto de Faria, Ana Carolina de Oliveira Tavares, Eva Mendes, Thaís Sales Antunes, Junio Cesar Jacinto De Paula e Denise Sobral
Epamig/Ilct
 Foto Divulgação Site Freepik
Foto Divulgação Site Freepik A mastite bovina, também conhecida como mamite, é uma inflamação da glândula mamária em vacas, sendo uma das principais doenças que afetam rebanhos leiteiros. Ela tem um impacto econômico significativo, resultando em queda na produção e qualidade do leite, além do descarte de vacas devido à perda de quarto(s) mamário(s) e custos adicionais de produção. Além disso, a mastite é relevante para a saúde pública, pois algumas bactérias envolvidas podem ser perigosas para os seres humanos. Esta doença resulta da infecção da glândula mamária, principalmente por bactérias do gênero Staphylococcus. Dentro deste gênero, Staphylococcus aureus é reconhecido como o principal patógeno da mastite bovina e seus principais sítios de localização nos animais parecem ser os quartos mamários infectados.
Diversos fatores de virulência podem ser produzidos por Staphylococcus aureus que contribuem para que a bactéria se defenda das formas fagocíticas do hospedeiro, facilite sua aderência às células epiteliais e a colonização dos tecidos, favorecendo sua persistência extracelular e garantindo, com êxito, sua instalação e manutenção nos tecidos do hospedeiro. Entre estes fatores está a produção de um mucopolissacarídeo extracelular (slime) que auxilia na aderência e na colonização do epitélio glandular mamário. A habilidade de S. aureus de aderir à superfície do epitélio tem sido associada à produção de biofilmes, que são descritos como aglomerações de células embebidas em matriz heterogênea extracelular, resultando em estruturas tridimensionais com características fisiológicas específicas (GAD et al., 2009).
Alguns polissacarídeos compõem o slime, mas um polissacarídeo específico de alto peso molecular, que tem a mesma função da cápsula bacteriana e intervém na aderência inicial das bactérias às superfícies dos polímeros, é denominado polissacarídeo capsular/adesina (PS/A) (GÖTZ, 2002).
O PS/A é descrito como componente da superfície celular e da camada do biofilme que protege as bactérias das defesas do hospedeiro e da fagocitose. Ele está envolvido no primeiro passo da adesão primária, que é seguida pela proliferação das células em agrupamentos de multicamadas (ARCIOLA; BALDASSARI; MONTANARO, 2001).
Cepas de S. aureus produzem Polissacarídeo Capsular (PC) in vivo ou sob condições culturais definidas. Estas cepas capsuladas são mais resistentes à absorção fagocítica do que as cepas não capsuladas (THAKKER et al., 1998). Em modelos de infecção estafilocócica em roedores, anticorpos anticapsulares protegeram animais contra a morte, a bacteremia, a endocardite, e a metástase para baço, fígado e rins (LEE et al., 1997).
Onze tipos de PC demonstrados por precipitação e aglutinação com antisoros monoespecíficos (SOMPOLINSKY et al., 1985). Há um consenso em relação ao fato de que PC5 e PC8 são os sorotipos predominantes em infecções estafilocócicas humanas. A avaliação da produção de cápsula por cepas de S. aureus isoladas de ruminantes mostrou resultados variados.
O presente estudo oferece insights valiosos sobre a caracterização fenotípica e molecular do Staphylococcus aureus em contextos de mastite bovina, contribuindo para a saúde e produtividade do rebanho leiteiro brasileiro.
Mastite bovina
Na bovinocultura leiteira, a mastite bovina é uma preocupação significativa para os produtores. Embora a forma clínica da mastite seja mais visível e cause apreensão, a forma subclínica é ainda mais comum e responsável por grandes prejuízos. Alguns especialistas preferem chamá-la de infecção subclínica. Algumas características importantes da mastite subclínica: ausência de sintomas visíveis; testes essenciais e prevalência silenciosa.
A mastite subclínica é um desafio porque não é facilmente identificada a olho nu. No entanto, seu impacto na produção de leite e nos custos de produção é significativo. Portanto, a detecção precoce e medidas de controle são essenciais para manter a saúde do rebanho e a qualidade do leite.
A mastite pode ter como causa diversos patógenos, mas são as bactérias os principais agentes etiológicos, normalmente divididas em duas categorias, os designados “contagiosos” e os “ambientais”. Na mastite ambiental, o reservatório do patógeno é o próprio ambiente, que pode estar presente no ar, cama, água e nas fezes das vacas leiteiras. Dentre os principais patógenos contagiosos encontra-se S. aureus. Os patógenos ambientais mais comuns são divididos em dois grupos, coliformes e estreptococos do ambiente (NATIONAL MASTITIS COUNCIL, 2001).
Staphylococcus aureus
A parede celular do S. aureus é composta por uma camada espessa de peptideoglicano (também conhecido como mureína). O peptideoglicano fornece resistência e forma à célula. Amembrana celular é uma estrutura bilaminar convencional, apresentando, em sua camada externa, ácidos lipoteicoicos ligados por pontes de dissacarídeos a um glicolipídeo. Esta espécie se caracteriza também por apresentar múltiplos fatores de virulência que contribuem para o estabelecimento e a manutenção da infecção (PEACOCK et al., 2002).
Para alcançar a persistência intracelular, S. aureus deve evitar a resposta imune e inflamatória do hospedeiro. Esta fina teia regulatória seria a chave da patogênese da infecção por S. aureus que conduz à cronicidade da doença e que, ao mesmo tempo, permite a adaptação do microrganismo a mudanças do meio ambiente durante o curso da infecção, bem como a sobrevivência e a persistência intracelular (TUCHSCHERR et al., 2010).
Staphylococcus aureus expressa também, em sua superfície, adesinas, proteínas antiopsonizantes (proteína A, fator de aglutinação A) e camada extracelular de polissacarídeos que impedem a fagocitose do mesmo (FOSTER, 2005). Na glândula mamária bovina, uma vez que o patógeno invade o órgão, ele deve superar a ação expulsiva da ordenha frequente. É por isso que adesão, sobrevivência e multiplicação de S. aureus no epitélio mamário são os primeiros eventos decisivos na patogênese da infecção. Este comportamento protegeria o patógeno da resposta imune do hospedeiro, do tratamento com antibióticos, e contribuiria para a sua persistência no tecido mamário (HEBERT et al., 2001).
Formação de biofilme
Diversos fatores de virulência, como exotoxinas, proteínas de superfície e polissacarídeos extracelulares de S. aureus, têm sido relatados em amostras isoladas de mastite bovina. Além disso, tem-se determinado que a formação de biofilme por estas cepas torna-se também um importante fator de virulência que contribui para a sua patogênese (TÜRKYILMAZ; ESKIIZMIRLILER, 2006).
A adesão de S. aureus ao epitélio da glândula mamária é considerada o primeiro ponto crítico na patogenia da mastite, sendo a maioria das cepas de S. aureus causadoras da doença circundada por uma camada polissacarídica espessa (slime) (VASUDEVAN et al., 2003).
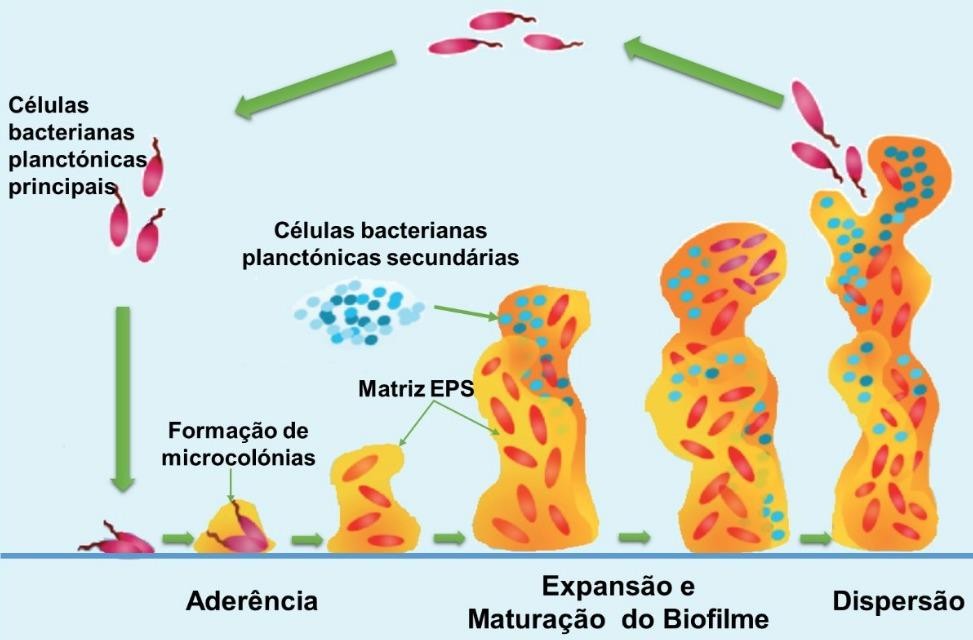
A habilidade de S. aureus de aderir à superfície do epitélio está associada à produção de biofilmes, composto de multicamadas de células embebidas em uma matriz polimérica extracelular (MELO et al., 2012) que exibem alteração fenotípica em relação ao crescimento planctônico (Figura 1).
Figura 1 Ciclo de desenvolvimento do biofilme (FOOD, 2024)
No biofilme, as bactérias são menos suscetíveis ao tratamento por antibióticos e à ação do sistema imune inato do hospedeiro. A capacidade de formação de biofilmes é fonte de estudo, tanto em medicina humana quanto veterinária, e a melhor compreensão desta característica fornece subsídios para a aplicação de medidas de tratamento mais eficazes na mastite bovina. A camada slime dificulta a fagocitose por células do sistema imunológico do hospedeiro, pois este mucopolissacarídeo facilita a adesão bacteriana a biomateriais, o qual não é removível, mesmo após lavagens sucessivas (DEGO; DIJK; NEDERBRAGT, 2002).
Sauer, Rickard e Davies (2007) identificaram biofilme como sendo um agrupamento de células microbianas associadas a superfícies, o qual se encontra envoltos por substâncias poliméricas extracelulares (Extracellular Polymeric Substances, EPS) hidratadas.
Diversos genes podem estar envolvidos na capacidade individual de cada cepa em formar biofilmes. Dentre estes, podem-se citar o gene ica (adessão intercelular) e o bap (proteína assoaciada ao biofilme), entre outros (KOZITSKAYA et al., 2004).
A matriz de EPS é responsável pela estrutura, coesão e integridade funcional do biofilme. Sua composição química, heterogênea e complexa (PEREIRA, 2001). Polissacarídeos, proteínas, fosfolipídeos, ácido teicóico e, até mesmo, ácidos nucleicos constituem as EPS (DONLAN; COSTERTON, 2002). O DNA extracelular, liberado pela autólise de células bacterianas, forma importante parte da matriz de EPS, influenciando tanto a estrutura do biofilme como a adesão inicial célula-superfície e célula-célula (THEERTHANKAR et al., 2010). No entanto, proteínas e polissacarídeos, que correspondem de 75% a 89% da composição das EPS, são os principais componentes (TSUNEDA et al., 2003).
Dentro deste grupo de genes, foi relatado que icaA e icaD desempenham papel importante na formação de biofilme em S. aureus. O gene icaA contém uma sequência sinal típica que codifica N-acetilglicosaminil transferase, uma enzima envolvida na síntese de oligômeros de N-acetilglicosamina a partir de UDP-N-acetilglicosamina (ARCIOLA; BALDASSARI; MONTANARO, 2001). Além disso, icaD tem sido associado à expressão de N-acetilglicosaminil transferase, resultando na expressão fenotípica do PC (GERKE et al., 1998).
Outros componentes, como ácidos teicoicos, proteínas de bactérias, incluindo proteína estafilocócica de superfície (staphylococcal surface protein-I-SSP-I), clumpling factor A, proteínas associadas ao biofilme (biofilm associated proteins-Bap) e DNA extracelular contribuem também para a estrutura do biofilme (VAUTOR et al., 2008).
Um dos mecanismos relacionados à produção de biofilme em S. aureus pode ser devido à existência da proteína Bap, sendo esta essencial para a fixação primária e a acumulação celular (SCARAMELLI; GONZÁLEZ, 2024).
O gene bap foi identificado em isolados de S. aureus de mastite bovina, nos quais tem papel fundamental na adesão em superfícies de poliestireno, adesão intercelular e formação de biofilme (EIFF; PETERS; HEILMANN, 2002).
Salimena et al. (2014) obsevaram que as células de S. aureus aderiram à superfície de polipropileno a partir de 48 horas, e a adesão aumentou em pequena proporção até 240 horas.
Conforme relatado por Arcuri (2000), na cadeia de produção de alimentos há correlação positiva entre a falha nos procedimentos de higiene e a formação destes filmes bacterianos, pois, havendo condições, as células aderidas evoluem para microcolônias e, assim, posteriormente, para o biofilme maduro. Além disso, a obtenção higiênica do leite e o atendimento a demais itens que compõem as boas práticas de processamento de alimentos são imprescindíveis para o controle destes microrganismos na cadeia alimentar.
Polissacarídeos capsulares
Staphylococcus aureus produzem polissacarídeos capsulares, tanto in vivo quanto em condições favoráveis de cultivo (LEE et al., 1993). Polissacarídeos capsulares (PC) são importantes na patogênese de infecções estafilocócicas e têm sido postulados como um dos principais fatores de virulência da bactéria (O'RIORDAN; LEE, 2004).
Foi proposta a existência de 11 sorotipos de PC isolados a partir de infecções humanas, no entanto, apenas quatro tipos (PC1, PC2, PC5 e PC8) foram isolados e caracterizados quimicamente, e não há evidência suficiente para concluir que os demais apresentem cápsulas ou estruturas quimicamente diferentes dos anteriores. Posteriormente, foi descrita a presença de outro tipo capsular, 336, em S. aureus isolados de mastite bovina; no entanto, sua estrutura química não foi caracterizada e demonstrou-se, recentemente, que os isolados que expressaram o polissacarídeo de superfície 336 tinham os genes cap5 ou cap8 (O’RIORDAN; LEE, 2004).
Existem diferentes métodos para a caracterização das cepas de S. aureus de acordo com o tipo de PC, e um dos mais utilizados consiste na utilização de anticorpos policlonais ou monoclonais específicos para as cápsulas dos tipos 1, 2, 5 e 8 (KARAKAWA et al., 1988). As cepas que não reagem com nenhum desses anticorpos são consideradas como não tipáveis (NT) porque os protótipos dessas linhagens não apresentam seus antissoros correspondentes (TUCHSCHERR et al., 2007). Cepas NT podem ser isoladas a partir 20% a 25% das infecções humanas (MIDDLETON; LUBY; ADAMS, 2009).
Amostras de S. aureus que apresentam os sorotipos 1 e 2 são altamente encapsuladas e originam colônias mucoides em meio sólido, facilmente observadas por microscopia. Os sorotipos 5 e 8 são denominados microencapsulados, já que produzem pouco material capsular, originando colônias não mucoides, compactas em meio sólido, que são indistinguíveis das colônias geradas por cepas não encapsuladas (O'RIORDAN; LEE, 2004).
De todos os sorotipos relatados, PC1 e PC2 são raramente encontrados, enquanto PC5 e PC8 foram presentes a partir de infecções humanas e bovinas. PC5 e PC8 são os tipos capsulares predominantes, presentes em 85%-90% dos isolados clínicos de S. aureus em diferentes estudos (ROGHMANN et al., 2005).
Estruturalmente, PC5 e PC8 demonstram grandes semelhanças. Ambos são compostos pelos mesmos açúcares. No entanto, diferem em algumas ligações, na configuração anomérica de um dos resíduos FucNAc e na localização do grupo O-acetil (JONES, 2005). Além disso, o locus do PC5 e PC8 é alélico e compreende uma região de 17,5 kb do cromossoma (O'RIORDAN; LEE, 2004), cada uma contendo 16 genes estritamente relacionados, a partir de capA a capP, transcritos em uma orientação (SAU et al., 1997). Por conseguinte, as sequências de aminoácidos previstas de 12 dos 16 genes de quadros de leitura abertos do grupo de genes cap5 e cap8 são quase idênticas. Os genes específicos se encontram na região central do locus que compreende os genes cap5H (cap8H) para cap5K (cap8K) (O'RIORDAN; LEE, 2004).
São descritas variações entre as amostras PC5 e PC8 de S. aureus isoladas de infecção intramamária bovina de diferentes localidades geográficas (REVELLI; RODRÍGUEZ, 2001).
Dois estudos realizados com S. aureus isolados de rebanhos leiteiros dos EUA (TOLLERSRUD et al., 2000) reportaram em torno de 40% de amostras tipáveis para os tipos capsulares 5 e 8, com prevalência do sorotipo 8. No estudo de Guidry et al. (1998), as amostras NT para os sorotipos 5 e 8 (59%) foram tipáveis para o sorotipo 336.
Na França, 69% dos 212 isolados recuperados a partir de leite de vacas eram do sorotipo 5 (51%) ou 8 (18%) (POUTREL et al., 1988). Em contrapartida, apenas 17% das amostras de S. aureus isoladas de mastite bovina em Israel produziram o sorotipo 5 ou 8 (SOMPOLINSKY et al., 1985). Em estudo realizado por Sordelli et al. (2000), apenas 14% das 195 estirpes bovinas isoladas na Argentina reagiram com anticorpos para PC5 ou PC8.
Staphylococcus aureus é um patógeno bacteriano relevante na mastite bovina. Produz vários fatores que contribuem para sua virulência. Esses componentes permitem que a bactéria se adira às células hospedeiras, cause danos aos tecidos e evite a resposta imunológica. O biofilme consiste na aglomeração de DNA extracelular, proteínas e polissacarídeos. A formação de biofilme favorece a adesão bacteriana e aumenta a atividade inflamatória. Estudos descreveram variações na prevalência de PC5 e PC8 entre amostras de S. aureus isoladas de infecções intramamárias bovinas em diferentes localidades geográficas. No entanto, nenhum estudo demonstrou a prevalência específica de algum sorotipo no Brasil. Essa informação seria fundamental para o desenvolvimento de estratégias de controle eficazes contra a mastite bovina no país.
Agradecimentos
Os autores agradecem o apoio da Fundação de Amparo à Pesquisa do Estado de Minas Gerais – FAPEMIG.
Autores:
Alessandra Pereira Sant’Anna Salimena, Bolsista de Pesquisa Nível I na Empresa de Pesquisa Agropecuária de Minas Gerais (EPAMIG) - Instituto de Laticínios Cândido Tostes (ILCT).
Letícia Scafutto de Faria, Bolsista de Pesquisa Nível I na Empresa de Pesquisa Agropecuária de Minas Gerais (EPAMIG) - Instituto de Laticínios Cândido Tostes (ILCT).
Ana Carolina de Oliveira Tavares, Bolsista de Pesquisa Nível III na Empresa de Pesquisa Agropecuária de Minas Gerais (EPAMIG) - Instituto de Laticínios Cândido Tostes (ILCT).
Eva Mendes, Mestranda em Ciência e Tecnologia do Leite e Derivados, Departamento de Farmácia UFJF e Técnica de Campo do programa ATeG na Bovinocultura de Leite - Sistema FAEMG SENAR.
Thaís Sales Antunes, Mestranda em Ciência e Tecnologia do Leite e Derivados, Departamento de Farmácia UFJF e Consultora em Laticínios, Técnica de Campo do programa de ATeG para Agroindústrias produtoras de Derivados Lácteos – Sistema FAEMG SENAR.
Junio Cesar Jacinto De Paula, Engenheiro de Alimentos, DSc. Professor e Pesquisador na Empresa de Pesquisa Agropecuária de Minas Gerais (EPAMIG) - Instituto de Laticínios Cândido Tostes (ILCT).
Denise Sobral, Engenheira de Alimentos, DSc. Professora e Pesquisadora da Empresa de Pesquisa Agropecuária de Minas Gerais (EPAMIG) - Instituto de Laticínios Cândido Tostes (ILCT).
Referências bibiolgráficas
ARCIOLA, C. R.; BALDASSARI, L.; MONTANARO, L. Presence of icaA and icaD and slime production in a collection of staphylococcal strains from catheter-associated infections. Journal of Clinical Microbiology, Washington, v. 39, n. 6, p. 2151-2156, Feb. 2001.
ARCURI, E. F. Biofilmes bacterianos na indústria de alimentos. Revista Leite e Derivados, São Paulo, v. 9, n. 53, p. 40-45, 2000.
DEGO, K. O.; DIJK, J. E. van; NEDERBRAGT, H. Factors involved in the early pathogenesis of bovine Staphylococcus aureus mastitis with emphasis on bacterial adhesion and invasion: a review. Veterinary Quarterly, The Hague, v. 24, n. 4, p. 181-198, Nov. 2002.
DONLAN, R. M.; COSTERTON, J. W. Biofilm: survival mechanisms of clinically relevant microorganisms. Clinical Microbiology Reviews, Washington, v. 15, n. 2, p. 167-193, Apr. 2002.
EIFF, C. von; PETERS, G.; HEILMANN, C. Pathogenesis of infections due to coagulase-negative staphylococci. Lancet Infectious Diseases, London, v. 2, n. 11, p. 677-685, Oct. 2002.
FOOD S. B. Biofilmes nas indústrias de alimentos: o que são e como se formam? Disponível em: https://foodsafetybrazil.org/biofilmes-o-que-sao-e-como-se-formam/. Acesso em: 12 Mar. 2024.
FOSTER, T. J. Immune evasion by staphylococci. Nature Reviews Microbiology, London, v. 3, n. 12, p. 948-58, Dec. 2005.
GAD, G. F. M. et al. Detection of icaA, icaD genes and biofilm production by Staphylococcus aureus and Staphylococcus epidermidis isolated from urinary tract catheterized patients. Journal of Infection in Developing Countries, Rome, v. 3, n. 5, p. 342-351, Mar. 2009.
GERKE, C. et al. Characterization of the N- acetylglucosaminyl-transferase activity involved in the biosynthesis of the Staphylococcus epidermidis - polysaccharide intercellular adhesin. Journal of Biology Chemesty, La Jolla, v. 273, n. 29, p. 18586-18596, 1998.
GÖTZ, F. Staphylococcus and biofilms. Molecular Microbiology, Baltimore, v. 43, n. 6, p. 1367-1378, 2002.
GUIDRY, A. J. et al. Serotyping scheme for Staphylococcus aureus isolated from cows with mastitis. American Journal of Veterinary Research, Beltsville, v. 59, n. 12, p. 1537-1539, Dec. 1998.
HERBERT, S. et al. Regulation of Staphylococcus aureus type 5 and type 8 capsular polysaccharide by CO2. Journal of Bacteriology, Washington, v. 183, n. 15, p. 4609-4613, Aug. 2001.
JONES, C. Revised structures for the capsular polysaccharides from Staphylococcus aureus Types 5 and 8, components of novel glycoconjugate vaccines. Carbohydrate Research, Amsterdam, v. 340, n. 6, p. 1097-1106, May 2005.
KARAKAWA, W. W. A. et al. Capsular antibodies induce type-specific phagocytosis of capsulated Staphylococcus aureus by human polymorphonuclear leukocytes. Infection and Immunity, Washington, v. 56, n. 5, p. 1090-1095, May 1988.
KOZITSKAYA, S. et al. The bacterial insertion sequence element IS256 occurs preferentially in nosocomial Staphylococcus epidermidis isolates: association with biofilm formation and resistence to aminoglycosides. Infection and Immunity, Washington, v. 72, n. 2, p. 1210-1215, Feb. 2004.
LEE, J. C. et al. Effects of in vitro and in vivo growth conditions on expression of type 8 capsular polysaccharide by Staphylococcus aureus. Infection and Immunity, Washington, v. 61, n. 5, p. 1853-1858, May 1993.
LEE, J. C. et al. Protective efficacy of antibodies to the Staphylococcus aureus type 5 capsular polysaccharide in a modified model of endocarditis in rats. Infection and Immunity, Washington, v. 65, n. 10, p. 4146-4151, July 1997.
MELCHIOR, M. B.; VAARKAMP, H.; FINK-GREMMELS, J. Biofilms: a role in recurrent mastitis infection? Veterinary Journal, Wageningen, v. 171, n. 3, p. 398-407, May 2006.
MELO, P. D. C. et al. Phenotypic and molecular analysis of biofilm production by Staphylococcus aureus strains isolated of bovine. Bioscience Journal, Uberlândia, v. 28, n. 1, p. 94-99, Jan./Feb. 2012.
MIDDLETON, J. R.; LUBY, C. D.; ADAMS, D. S. Efficacy of vaccination against staphylococcal mastitis: a review and new data. Veterinary Microbiology, Wageningen, v. 134, n. 1/2, p. 192-198, Feb. 2009.
NATIONAL MASTITIS COUNCIL. Current concepts of bovine mastitis. 4th ed. Madison, 2001. 64 p.
O'RIORDAN, K.; LEE, J. Staphylococcus aureus capsular polysaccharides. Clinical Microbiology Reviews, Washington, v. 17, n. 1, p. 218-234, Jan. 2004.
PEACOCK, S. J. et al. Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infection and Immunity, Washington, v. 70, n. 9, p. 4987-4996, Sept. 2002.
PEREIRA, M. O. P. O. Comparação da eficácia de dois biocidas (carbamato e glutaraldeído) em sistemas de biofilme. 2001. 234 p. Tese (Doutorado em Engenharia Química e Biológica) - Universidade do Minho, Braga, 2001.
POUTREL, B. et al. Prevalence of capsular polysaccharide types 5 and 8 among Staphylococcus aureus isolates from cow, goat, and ewe milk. Journal of Clinical Microbiology, Washington, v. 26, n. 1, p. 38-40, Jan. 1988.
REVELLI, G.; RODRÍGUEZ, C. G. Prevalencia de agentes etiológicos causales de mastitis bovina en la zona noroeste de Santa Fe y sur de Santiago del Estero: respuesta a la sensibilidad antimicrobiana. Tecnología Láctea Latinoamericana, Buenos Aires, v. 6, n. 23, p. 48-53, 2001.
ROGHMANN, M. et al. Epidemiology of capsular and surface polysaccharide in Staphylococcus aureus infections complicated by bacteraemia. Journal of Hospital Infection, Baltimore, v. 59, n. 1, p. 27-32, Jan. 2005.
SALIMENA, A. P. S. et al. Scanning electron microscopy of biofilm formation by Staphylococcus aureus on stainless steel and polypropylene surfaces. African Journal of Microbiology Research, Nairobi, v. 8, n. 34, p. 3136-3143, Aug. 2014.
SAU, S. et al. The Staphylococcus aureus allelic genetic loci for serotype 5 and 8 capsule expression contain the type-specific genes flanked by common genes. Microbiology, New York, v. 143, n. 7, p. 2395-2405, July 1997.
SAUER, K.; RICKARD, A. H.; DAVIES, D. G. Biofilms and biocomplexity. Microbe-American Society for Microbiology, Washington, v. 2, n. 7, p. 347-353, 2007.
SCARAMELLI, A.; GONZÁLEZ, Z. Epizootiología y diagnóstico de la mastitis bovina.
Disponível em: . Acesso em: 12 Mar. 2024.
SOMPOLINSKY, D. et al. Encapsulation and capsular types in isolates of Staphylococcus aureus from different sources and relationship to phage types. Journal of Clinical Microbiology, Washington, v. 22, n. 5, p. 828-834, Nov. 1985.
THAKKER, M. et al. Staphylococcus aureus serotype 5 capsular polysaccharide is antiphagocytic and enhances bacterial virulence in a murine bacteremia model. Infection and Immunity, Washington, v. 66, n. 11, p. 5183-5189, Nov. 1998.
THEERTHANKAR, D. et al. Role of extracellular DNA in initial bacterial adhesion and surface aggregation. Applied and Environmental Microbiology, Washington, v. 76, n. 10, p. 3405-3408, May 2010.
TOLLERSRUD, T. et al. Genetic and serologic evaluation of capsule production by bovine mammary isolates of Staphylococcus aureus and others Staphylococcus spp. from Europe and the United States. Journal of Clinical Microbiology, Washington, v. 38, n. 8, p. 2998-3003, Aug. 2000.
TSUNEDA, S. et al. Extracellular polymeric substances responsible for bacterial adhesion onto solid surface. FEMS Microbiology Letters, Amsterdam, v. 223, n. 2, p. 287-292, June 2003.
TUCHSCHERR, L. et al. Staphylococcus aureus adaptation to the host and persistence: role of loss of capsular polysaccharide expression. Future Microbiology, London, v. 5, n. 12, p. 1823-1832, Dec. 2010.
TUCHSCHERR, L. P. N. et al. Characterization of a new variant of IS257 that has displaced the capsule genes within bovine isolates of Staphylococcus aureus. Infection and Immunity, Washington, v. 75, n. 11, p. 5483-5488, Nov. 2007.
TÜRKYILMAZ, S.; ESKIIZMIRLILER, S. Detection of slime factor production and antibiotic resistance in Staphylococcus strains isolated from various animal clinical samples. Turkish Journal of Veterinary and Animal Sciences, Ankara, v. 30, n. 2, p. 201-206, Apr. 2006.
VASUDEVAN, P. et al. Phenotypic and genotypic characterization of bovine mastitis isolates of Staphylococcus aureus for biofilm formation. Veterinary Microbiology, Wageningen, v. 92, n. 1/2, p. 179-185, Mar. 2003.
VAUTOR, E. et al. Evaluation of the presence of the bap gene in Staphylococcus aureus isolates recovered from human and animalsspecies. Veterinary Microbiology, Wageningen, v. 127, n. 3, p. 407-411, Mar. 2008.
Fonte: Empresa de Pesquisa Agropecuária de Minas Gerais/Instituto de Laticínios Cândido Tostes

 Foto Divulgação Site Freepik
Foto Divulgação Site Freepik